consideraÇÕes
gerais sobre os
mecanismos de “feedback”
Bernadete
M.V. Amin
1 - “feedback” por
ações hormonais
diversas
A
regulação das funções
corporais pelos
hormônios é feita
pela regulação das
atividades
das células individuais.
Os hormônios sinalizam
às células para que
comecem ou
parem de secretar, que
se contraiam, se dividam ou se
diferenciem. Também
podem acelerar ou
tornar mais
lentos esses
processos, ou
podem modificar as respostas
a outros hormônios.
Todas essas ações
celulares
se somam para produzir
as respostas biológicas que observamos ao nível
dos
tecidos, dos órgãos
e do corpo todo.
Apesar dos hormônios
se distribuírem por todo
o sangue e por
todo o líquido
extracelular, apenas certas células-alvo respondem a um
dado hormônio.
Algumas células são
alvos de mais
de um hormônio.
Se
uma célula responde ou não
a um hormônio
depende de ter ou
não receptores para aquele hormônio.
Entretanto
é importante reconhecer
que a resposta
específica provocada em uma célula
é
determinada mais
pela própria
célula que
pelo hormônio,
e que tipos
celulares diferentes
podem responder ao mesmo
estímulo de maneiras
diferentes. Por
exemplo, tanto
as células musculares
lisas vasculares, quanto
as células dos ductos
coletores dos túbulos renais, são
alvos do hormônio
da hipófise posterior, a
vasopressina. Quando
estimuladas pela vasopressina,
as arteríolas se contraem, enquanto
os ductos
coletores aumentam sua
permeabilidade à água.
A manutenção da integridade do ambiente
interno, ou
o enfrentamento bem-sucedido
de um desafio
externo, tipicamente implicam a interação coordenada
de vários sistemas
fisiológicos e a integração
de múltiplos sinais
hormonais. As soluções
dos problemas
fisiológicos requerem a integração
de grande variedade
de eventos simultâneos
que, em
conjunto, podem produzir
resultados que
são maiores
ou menores
que a simples
soma algébrica das respostas
hormonais individuais.
2 - “feedback” por
modulação
Nem todos
os aspectos do controle
hormonal são determinados
simplesmente
pela quantidade
de hormônio secretada, nem
mesmo pelo
momento em
que o hormônio
é secretado.
A receptividade dos tecidos-alvos à estimulação
hormonal não
é constante e pode alterar-se
sob várias circunstâncias.
A receptividade dos tecidos-alvo à estimulação
hormonal pode ser
expressada em termos
de dois aspectos
distintos, mas
relacionados: a sensibilidade
à
estimulação e a capacidade
de resposta. A sensibilidade
descreve a acuidade da capacidade de uma célula
reconhecer um
sinal e de responder
em proporção
à intensidade daquele sinal. Podemos definir
sensibilidade em
termos da concentração
do hormônio que
provocará cerca de 50%
da resposta máxima.
A capacidade de resposta,
ou a resposta
máxima que
um tecido
é capaz de apresentar depende
da quantidade de células
componentes ou
diferenciadas naquele tecido, bem como do
nível de desenvolvimento
da maquinaria enzimática
naquelas células.Os hormônios
regulam tanto a sensibilidade
quanto a capacidade
dos tecidos-alvos responderem a estes
ou a outros
hormônios.
3 - “feedback” via
receptores
Um mecanismo
pelo qual
os hormônios determinam a
sensibilidade
dos tecidos-alvo é a regulação dos receptores
do hormônio. Deve ser
lembrado que o evento
inicial na produção
de uma resposta hormonal
é a interação
do hormônio com
seu receptor.
Quanto
maior a concentração
do hormônio, maior
a probabilidade de ocorrer
interação com
seus receptores.
Se não existirem receptores
do hormônio, no entanto,
não pode haver
resposta, e quanto
mais receptores
disponíveis para
interagir com
uma dada quantidade
de hormônio, maior
a probabilidade de haver
resposta. Em
outras palavras, a probabilidade
da ocorrência da interação
hormônio-receptor está relacionada com
a
abundância do hormônio
e
do receptor. Apesar
da afinidade do receptor
por seu
hormônio também
poder ser
modulada, de modo geral
parece ser o número
de receptores, em
vez de sua
afinidade pelo
hormônio, que
é usualmente
modulado.
Alguns hormônios
diminuem o número de seus próprios
receptores nos
tecidos-alvos. Este fenômeno,
chamado de “regulação para baixo”
(down regulation), foi reconhecido como
verdadeiro na endocrinologia
moderna, quando
se mostrou que a sensibilidade
diminuída de algumas células
à insulina, nos estados
hiperinsulínicos, resultava
da diminuição do número
de receptores na superfície
da célula. No entanto,
um fenômeno
semelhante foi observado,
muitos anos
antes, por
Cannon e Rosenblueth, que
descreveram a
“supersensibilidade dos tecidos
desnervados”. A descoberta original desse fenômeno
por Cannon dizia respeito
à hipersensibilidade do coração
desnervado
à epinefrina e norepinefrina circulantes. A generalização
deste fenômeno em
ambos os sistemas
de controle endócrino
e neural é indicada adicionalmente
pelo aumento dos
receptores da acetilcolina que
ocorre após um
músculo ser
desnervado e da restauração
à
normalidade após a
reinervação. O fenômeno
da taquifilaxia, ou perda
da capacidade de resposta
a um agente
farmacológico, após
exposição repetida ou
constante, pode ser
outro exemplo
da regulação para baixo
dos receptores. A
regulação para baixo pode resultar
da inativação dos receptores
na superfície da célula,
da destruição
aumentada dos complexos
hormônio-receptor internalizados, ou
da síntese
diminuída dos receptores.
A
regulação para baixo
não é limitada aos
efeitos
de um hormônio
sobre seu
próprio receptor,
ou aos receptores
de superfície para
hormônios
hidrossolúveis. Um hormônio pode regular
para baixo
receptores para
outro hormônio.
Este parece ser
o mecanismo pelo
qual T3 diminui a sensibilidade
dos tireotrofos da hipófise
ao hormônio de liberação
da tireotrofina. De modo semelhante, a progesterona
pode regular para
baixo seu
próprio receptores
e também o do estrogênio.
A
regulação para cima
(up reguiation), ou o aumento
dos receptores disponíveis,
também ocorre e pode ser
vista tanto
nos receptores
de superfície. quanto
nos receptores
internos dos hormônios
lipossolúveis. A prolactina e possivelmente o hormônio
do crescimento podem regular para cima seus receptores
nas células
que respondem. O estrogênio
regula para cima
tanto seus
próprios receptores.
quanto os do hormônio
luteinizante e do hormônio
folículo-estimulante nas células
do ovário,
durante o ciclo
menstrual. Os mecanismos
moleculares da
regulação para cima
não são
compreendidos, atualmente, e são o assunto
de pesquisas intensas. Parece que a regulação para cima, tal como
a produzida pelo estrogênio,
é iniciada
por alteração da expressão
dos genes.
A sensibilidade à
estimulação
hormonal também pode ser
modulada de modo que
não envolva os receptores.
A modulação
pós-receptor pode afetar qualquer uma
das etapas da via
biológica pela qual
são produzidos os efeitos
hormonais. Por exemplo,
a atividade da fosfodiesterase
do
monofosfato cíclico de adenosina (AMPc.) aumenta
no tecido adiposo na ausência dos hormônios
hipofisános. Deve ser lembrado que
essa enzima catalisa a degradação do AMPc. E, quando
sua atividade
está aumentada. menos
AMPc pode
acumular-se, após a
estimulação da adenilil ciclase por
um hormônio,
como a
epinefrina. Portanto, se todas
as outras
condições forem iguais,
seria necessária concentração
mais alta
de epinefrina para produzir
uma dada quantidade
de lipólise do que
aquela necessária em presença
de quantidades
normais de hormônios
hipofisários e daí a sensibilidade
à
epinefrina parecer reduzida.
A atividade aumentada da
fosfodiesterase é apenas
um dentre
vá rios fatores que
contribuem para a sensibilidade
diminuída à epinefrina no tecido
adiposo
de animais
hipo-hipofisários. Esses
tecidos também
têm um defeito na sua
capacidade de responder
ao AMPc por causa
de deficiências da lipase
sensível ao hormônio. Portanto,
mesmo
quando todos
os receptores estão ocupados
a resposta máxima
que esses
tecidos podem produzir
fica abaixo do normal.
4 - “feedback” via
potencialização
Os hormônios também
podem aumentar a capacidade
das células ou
tecidos-alvo responderem a outros
estímulos. Por exemplo,
o estradiol aumenta
a síntese de prolactina nos lactotropos hipofisários e, portanto,
aumenta a secreção
de prolactina em resposta
aos estímulos
fisiológicos. Modo ainda mais
óbvio pelo
qual um
hormônio pode aumentar
a capacidade
do tecido responder
a estímulos
fisiológicos é aumentar ou diminuir o número de células
nos
tecidos-alvo. Por exemplo,
o aumento gradativo
da secreção do
estradiol visto na parte inicial
do ciclo
menstrual, reflete a atividade
de número crescente
de células da granulosa,
em vez
de alteração da secreção
da
gonadotropina.
5 - “feedback” permissivo
Outro aspecto
da modulação
hormonal relacionado com os exemplos
acima foi chamado de ação
permissiva. Um
hormônio atua permissivamente
quando sua
presença é necessária
para, ou
permite, que ocorra uma resposta biológica, mesmo
apesar do hormônio
não iniciar
a resposta. Efeitos
permissivos foram originalmente
descritos para os hormônios
corticais adrenais, mas parecem
ocorrer também
para outros hormônios.
As ações
permissivas não são
limitadas às respostas
aos hormônios, mas
ocorrem em qualquer
resposta celular
a qualquer sinal.
Apesar de um
hormônio poder
desencadear uma resposta
global celular
afetando alguma reação
fundamental determinadora da intensidade
dessa resposta, mais
de um processo
pode ser afetado.
Os efeitos hormonais exercidos em vários
locais, em
uma mesma célula,
se somam para produzir
a resposta global.
Consideremos, por exemplo,
apenas algumas das maneiras
pelas quais a insulina
atua sobre a célula
adiposa para
promover o armazenamento de
triglicerídeos:
1.
Atua na membrana celular,
para aumentar a disponibilidade de substrato
para a síntese
de lipídios
2. Ativa várias enzimas
citosólicas e mitocondriais, críticas
para a síntese
de ácidos
graxos
3.
Inibe a degradação
dos
triglicerídeos já
formados
4.
Induz a síntese da enzima extracelular lipase lipoproteica, necessária à
captação de lipídios
da circulação
Qualquer um
desses efeitos poderia
cumprir o objetivo
de aumentar o armazenamento de gordura,
mas, coletivamente,
estes diferentes
efeitos tornam possível
uma faixa muito
mais ampla
de resposta, em
tempo mais
curto.
O reforço também
pode ser observado
ao nível de todo
o organismo, onde
um hormônio
pode atuar de maneiras
diferentes em
tecidos diversos
para produzir efeitos complementares.
Um bom
exemplo disso é a ação
dos hormônios
glicocorticóides promovendo
gliconeogênese. Eles
atuam nos tecidos periféricos,
para mobilizar substrato, e
no fígado, para
aumentar a conversão
dos precursores a glicose.
Tanto a ação
extra-hepática quanto a ação
hepática, aumentariam a
gliconeogênese, mas, em conjunto,
essas ações
complementares reforçam
uma à outra e aumentam a
magnitude
e a rapidez da resposta
global.
6 - “feedback” via
redundância
As funções cruciais
são governadas por
mecanismos à prova
de erro. Tal como
cada sistema
de órgãos
contém em
si um
excesso de capacidade,
que lhe
confere o potencial de funcionar
a níveis além
das demandas do dia-a-dia,
assim também
há excesso de capacidade
reguladora, fornecido sob a forma de controles
aparentemente duplicados ou
super-postos. Falando simplesmente, o corpo
tem mais de uma maneira
de atingir um
determinado objetivo.
Por exemplo
a conversão do
glicogênio hepático
a glicose
sanguínea pode ser
sinalizada por, pelo
menos, dois
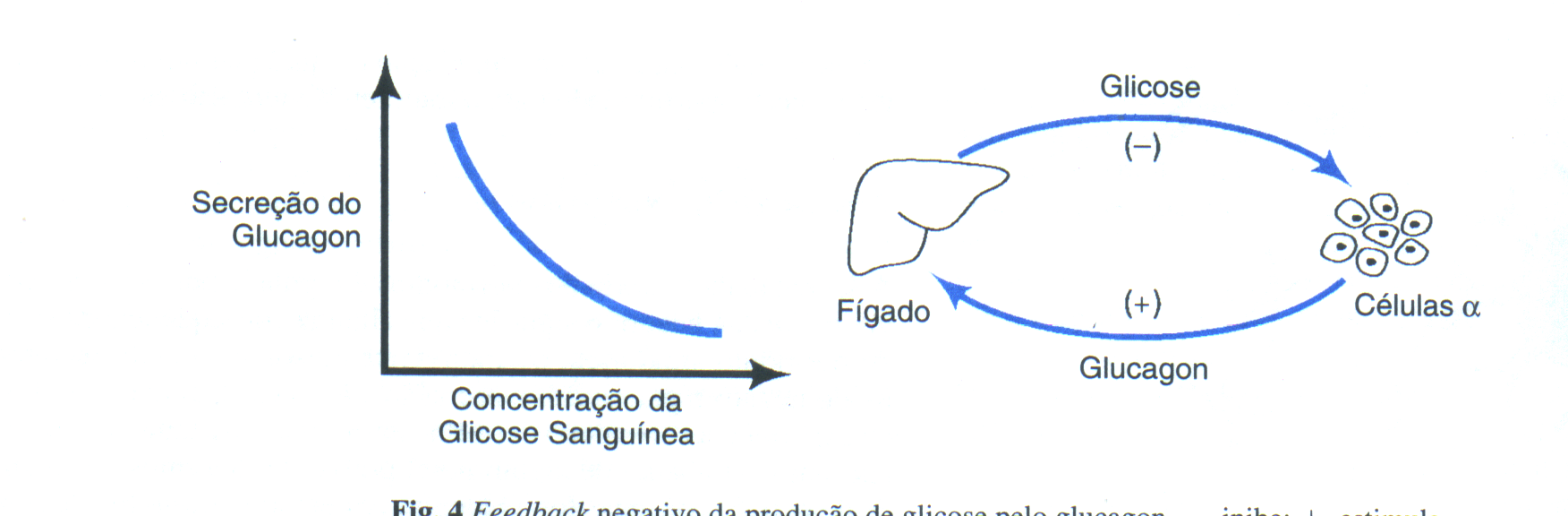
hormônios, o glucagon das células
alfa do pâncreas
e a epinefrina da medula
adrenal. Ambos esses hormônios
aumentam a produção
de AMPc no fígado e
desse modo, ativam a enzima
fosforilase, que catalisa a
glicogenólise. Dois hormônios
secretados, a partir de dois
tecidos diferentes,
às vezes em
resposta a diferentes
condições,
produzem assim o mesmo
resultado final.
A redundância também
pode ser vista
a nível molecular. Usando o mesmo
exemplo da conversão
do glicogênio hepático
a glicose sanguínea,
há, até mesmo,
duas maneiras pelas quais
a epinefrina pode ativar a fosforilase.
Estimulando
os receptores
beta-adrenérgicos, a
epinefrina aumenta a formação de AMPc como
já mencionado.
Estimulando os receptores alfa
adrenérgicos, a epinefrina também
ativa a fosforilase, mas
esses receptores
operam por meio
das concentrações
aumentadas de cálcio
intracelular
produzidas pela liberação
de inositol trifosfato.
Os mecanismos redundantes
não apenas
asseguram que um
processo crítico
irá ocorrer, mas
também oferecem a
possibilidade da
flexibilidade e do ajuste fino e sutil do processo. Apesar
de redundantes, quanto
a dois diferentes
hormônios poderem ter
alguns efeitos
superpostos, a ações
dos dois
hormônios usualmente
não são
idênticas em todos
o aspectos. Dentro
da faixa fisiológica de suas concentrações
no sangue as ações
do glucagon são
restritas ao fígado; a
epinefrina produz várias
outras respostas em
muitos tecidos
extra-hepáticos, enquanto
aumenta glicogenólise no fígado.
As variações da entrada
relativa de ambos
o hormônios permitem um amplo
espectro de alterações das concentrações
da glicose sanguínea,
em relação
a outros efeitos
da epinefrina, como o aumento
da freqüência cardíaca.
Dois hormônios
que produzem efeitos
comuns podem diferir
não apenas
na sua gama
de ações, mas
também quanto
a suas constantes de tempo.
Um deles pode ter
instalação mais
rápida e duração
mais
curta de sua
ação, enquanto
o outro pode ter
duração mais
longa da ação,
mas instalação
mais lenta.
Por exemplo,
a epinefrina aumenta as concentrações sanguíneas
dos ácidos
graxos livres
(AGL) dentro de segundos
a minutos, e esse
efeito se dissipa de modo igualmente
rápido,
quando cessa a secreção
de epinefrina. O hormônio do crescimento
também aumenta
as concentrações
sanguíneas dos AGL mas, seus efeitos
são vistos apenas
após período
de latência de 2 ou 3 horas e
persistem por muitas horas. Um
hormônio como
a
epinefrina pode, portanto, ser usado para atender
as necessidades a curto
prazo, e outro,
como o hormônio
do crescimento, pode responder
as necessidades prolongadas.
Uma
das implicações
da redundância, para a compreensão
tanto
da fisiologia normal
quanto das doenças
endócrinas, é que
a insuficiência
parcial, ou
talvez mesmo
completa, de um
mecanismo pode ser
compensada pelo
suporte aumentado de outro
mecanismo. Assim,
as deficiências funcionais
podem ser evidentes
apenas de modo
sutil e podem não
se apresentar abertamente
como uma doença.
Algumas deficiências
podem tornar-se aparentes apenas
após a provocação
apropriada ou a
perturbação do sistema.
Inversamente, estratégias
para intervenções
terapêuticas,
projetadas para aumentar
ou diminuir
a intensidade ou
a velocidade de um
processo, têm que
levar em
consideração as entradas
redundantes que
regulam esse processo,
A mera aceleração,
ou o bloqueio de uma entrada reguladora, pode não
produzir o efeito
desejado porque ajustes
independentes podem
compensar completamente a intervenção.
7 - “feedback” via
Mecanismos Empurra-Puxa
(Push: Pull)
Muitos processos
críticos estão sob
controle duplo
por agentes
que atuam antagonisticamente, quer para estimular,
quer para
inibir. Este
controle duplo
permite a
regulação mais precisa,
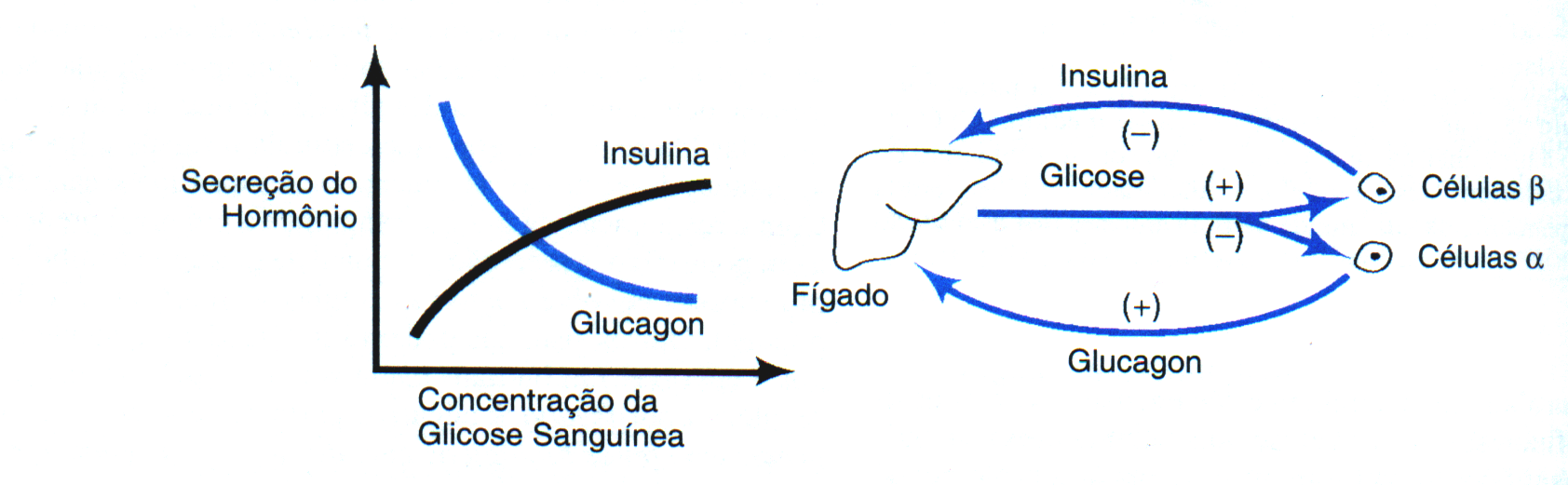
através do “feedback” negativo. O exemplo
citado anteriormente era a produção
hepática de glicose,
que é aumentada pelo
glucagon e inibida pela insulina. Em
situações de emergência,
ou durante
o exercício, a
epinefrina e
noraepinefrina, liberadas a partir da medula adrenal e das terminações
nervosas simpáticas, sobrepujam ambos
os
sistemas de “feedback” negativo, inibindo a secreção
de insulina e estimulando a secreção de glucagon (11). O efeito
de acrescentar uma influência
estimuladora e remover, simultaneamente,
uma
influencia inibitória é a obtenção da resposta
rápida e intensa,
mais rapida e maior
do que poderia
ser realizado afetando simplesmente
cada um
dos hormônios isoladamente, ou
a da que poderia
ser realizada pelo
efeito glicogenolítico direto
da epinefrina ou da
norepinefrina.
Outro tipo
de mecanismo empurra-puxa (push: pull) pode
ser visto
a nível molecular. A síntese
final do glicogênio, a partir
da glicose, depende da atividade de duas enzimas,
a glicogênio sintetase que
catalisa a formação
de glicogênio a partir
da glicose, e a
glicogênio fosfarilase, que
catalisa a degradação
do glicogênio- (Fig. l2). A velocidade
final da reação
é determinada pelo
balanço da atividade
das duas enzimas. A atividade
de ambas as enzimas está
sujeita à
regulação pela
fosforilação, mas
em
direções opostas:
a adição
de um grupamento
fosfato ativa
a fosforilase, mas inativa
a sintetase. Nesse caso, um único
agente, o AMPc, que
ativa a proteína
cinase A, aumenta a atividade
da fosforilase e inibe simultaneamente a sintetase.
Referência Bibliográfica
JOHNSON
Leonard R. Fundamentos
de Fisiologia Médica.
2 ed.Guanabara Koogan; Rio de Janeiro. 1998.